
Условия внешней среды, такие как температура, влажность, нали¬чие кислорода, освещенность, осмотическое и гидростатическое давление и другие факторы, влияют на рост микроорганизмов, распространение их в природе и поведение в технологических процессах.
Концентрация среды (осмотическое давление). Каждый организм способен расти и размножаться только при определенных концентрациях солей в среде. Изменение оптимальной концентрации среды приводит к ослаблению жизнедеятельности, гибели клеток. Осмотическое давление возникает из-за стремления воды проникнуть через полупроницаемую мембрану в сторону более концентрированного из двух разделенных этой мембраной растворов. Такое давление прямо пропорционально концентрации молекул веществ, которые не могут пройти через мембрану.
Когда концентрация среды очень низкая, клетка испытывает гипоосмотический стресс и стремится впитать воду. В плотной среде – гиперосмотический стресс – клетка теряет воду и «сморщивается», то есть происходит плазмолиз, нарушается обмен веществ и клетка гибнет.
Степень стресса зависит от плотности среды и типа сахаров, а также физиологического состояния клеток. Растущие клетки более чувстви¬тельны к стрессу, чем клетки из стационарной фазы. В гипоосмотических условиях с целью противостоять поступлению воды в клетки происходит стимуляция синтеза клеточной стенки.
Для S. cerevisiae отмечено изменение размера и формы клеток с повышением концентрации сухих веществ в среде. Независимо от морфологических особенностей штаммов в плотных средах дрожже¬вые клетки становятся круглыми, кроме того, наблюдается увеличе¬ние плотности цитоплазмы. Изменяется также метаболизм углеводов: увеличивается синтез глицерина.
Микроорганизмы, способные развиваться в среде с высоким осмо¬тическим давлением, т. е. устойчивые к высоким концентрациям соли и сахара в среде, называют осмофильными. Осмофильные штаммы дрожжей вызывают брожение меда при получении медовых напитков, а также используются в высокоплотном пивоварении.
Известны организмы, способные расти лишь при очень высоких концентрациях солей (NaCl). Это галофильные, т. е. «любящие» высо¬кую концентрацию солей, организмы.
Содержание влаги. Осмотическое давление в клетке определяется содержанием воды. Вода в этом случае играет механическую роль, поддерживая тургор клеток. Это одна из важнейших функций воды в живых организмах.
Кроме того, вода играет структурную роль, обеспечивая гидрата¬цию белков и других клеточных компонентов, она служит растворите¬лем для клеточных метаболитов, например аминокислот.
Вода также является химическим реагентом, участвуя в реакциях гидролиза и конденсации. Содержание свободной влаги в клетках составляет в среднем 75–85% и может меняться в зависимости от условий среды, в которых клетка находится. При потере воды клеткой происходит ее высыхание, нарушается обмен веществ, что в конце концов приводит к гибели клетки.
Микроорганизмы могут жить при различной влажности, но для каждого вида характерна своя оптимальная влажность. Так, мно¬гие мицелиальные грибы родов Aspergillus и Pénicillium могут расти на едва увлажненных субстратах. Известно, что при хранении зерна про¬исходит смена микробиоты ячменя. Количество полевых мицелиальных грибов на зерне уменьшается и заметно возрастает количество грибов хранения. Это связано с тем, что полевые грибы для своего развития требуют более высокой влажности (около 25%), а влажность ячменя, направляемого на хранение, составляет 14–15%. Грибы же хранения способны развиваться даже при влажности субстрата ниже 12–13%. К наиболее часто встречающимся грибам хранения отно¬сятся отдельные виды родов Pénicillium и Мисоr, представители рода Aspergillus – Asp. niger, Asp. flavus, Asp. fumigatus, Asp. clavatus, Asp. glaucus, a также Rhizopus nigricans.
Содержание воды в клетке выражается как осмотическое давление (в атм), тоничность среды (осмоли) или активность воды (άw).
Параметр «άw» обычно используется при изучении концентрированных сред, чаще всего при изучении порчи продуктов, которая тем меньше, чем меньше активность воды, которая в растворе выражается отношением давления водяного пара раствора к давлению чистой воды при той же температуре. Активность воды (άw) эквивалентна относи¬тельной влажности атмосферы в равновесии с раствором.
Ниже приведены показатели активности воды для некоторых групп микроорганизмов:
| Большинство бактерий | 0,95-0,91 |
| Большинство дрожжей, вызывающих порчу пищевых продуктов | 0,88-0,65 |
| Большинство плесневых грибов, вызывающих порчу пищевых продуктов | 0,80-0,60 |
| Грамположительные палочки | 0,97-0,96 |
| Грамположительные кокки | 0,90-0,86 |
| Галофильные бактерии | 0,75 |
| Ксерофильные плесневые грибы | 0,65 |
| Осмофильные дрожжи | 0,60 |
Низкую активность воды хорошо переносят осмофильные дрожжи, для которых минимальное значение активности воды 0,6.
В случае сред, применяемых для культивирования микроорганиз¬мов, активность воды мало отличается от единицы.
Механизм губительного действия высушивания используют для консервирования продуктов с целью воздействия на посторонние микроорганизмы. Например, патогенные нейссерии и многие другие бактерии погибают при высушивании в течение нескольких ми¬нут, шигеллы, сальмонеллы, холерный вибрион – в течение несколь¬ких суток, а микобактерии туберкулеза выдерживают высушивание более 3 мес. Максимальной устойчивостью к высушиванию обладают споры бактерий и мицелиальных грибов, которые сохраняют способ¬ность к прорастанию в течение 10 и 20 лет соответственно.
Температура. Микроорганизмы лишены механизмов, регули¬рующих температуру, поэтому их существование определяется тем¬пературой окружающей среды. Рост каждого вида микроорганиз¬мов может происходить в определенном температурном интервале, ограниченном минимальной и максимальной температурой. Наибо¬лее благоприятная для роста температура называется оптимальной. Для каждого микроорганизма оптимальная температура определяется ее влиянием на активность ферментов и множество ферментативных реакций, происходящих в клетке. С понижением температуры актив-ность большинства ферментов падает, однако этой зависимости под¬чиняются не все ферменты. Активность некоторых утрачивается уже при температуре минус 20°С. А некоторые, такие как каталаза и пероксидаза, активны даже в замороженном состоянии. В общем случае с повышением температуры активности ферментов увеличиваются и скорости биохимических реакций. Однако при увеличении темпе¬ратуры повышается и термическая денатурация, поэтому, пройдя оп¬тимум, активность ферментов падает.
Ферменты микроорганизмов имеют оптимумы температурной активности соответственно экологии их обитания. По отношению к температуре микроорганизмы могут быть разделены на группы (табл. 2).
Таблица 2
Распределение микроорганизмов по их отношению к температуре
| Группа | Температура, °С | ||
| Минимум | Оптимум | Максимум | |
|
Психрофилы: облигатные факультативные |
-5…+5 -5...+5 |
5-15 25-30 |
20-22 30-35 |
| Мезофилы | 10-15 | 30-45 | 35-47 |
|
Термофилы: облигатные факультативные термотолерантные |
40-42 20 20 |
65-70 50-65 45-50 |
75-80 55-75 50-70 |
Большинство почвенных и водных бактерий представлены мезофильной группой, их температурный оптимум лежит в пределах от 20 до 45 °С. Подавляющая часть патогенных микроорганизмов также являются мезофилами.
Некоторые микроорганизмы лучше всего растут при температуре выше 45°С (спорообразующие бактерии), их называют термофиль¬ными.
Другую крайность представляют психрофильные организмы. Это преимущественно морская микробиота (светящиеся бактерии) и же¬лезобактерии (Gallionella), для которых оптимум температуры менее 20 °С. Термотолерантными называют бактерии, у которых темпера¬турный оптимум лежит в области средних температур, но которые способны переносить и более высокие температуры, например Methylococius capsulatus: оптимальная температура – 37 °С, а макси¬мальная – 50 °С.
Ввиду того что скорость протекания реакций в дрожжевых клет¬ках прежде всего отражается на удельной скорости их роста, можно считать, что 1n μ = f(1/T). Очень ценным является показатель энергии активации, значение которого в пределах оптимальной температуры роста для каждого вида микроорганизма постоянно, но увеличивается при переходе к низким и высоким температурам. Показано, что тем-пературный диапазон роста микроскопических грибов, к которым от¬носятся и дрожжи, находится около (30,0 ± 3,0) °С.
Изменение температуры часто сопровождается изменениями по¬требности микроорганизмов в факторах роста, которые являются, как правило, активной частью ферментов.
Для S. cerevisiae установлено, что при температуре 38ºС потреб¬ность клеток в пантотеновой кислоте значительно превышает пот¬ребность дрожжей в этом витамине при 30ºС. Кроме того, положи¬тельный эффект наблюдается при внесении в среду хлорида натрия. Температура влияет на уровень РНК, белка, липидный и углеводный состав клеток; например, при одной и той же скорости роста содер¬жание РНК в дрожжах при снижении температуры увеличивается в несколько раз. Также в зависимости от температуры колеблется со¬держание белка в дрожжах, что будет коррелировать с природой ли¬митирующего субстрата. При снижении температуры увеличивается содержание ненасыщенных жирных кислот (ННЖК). Кроме этого, имеет значение скорость снижения температур и перепад температур во время брожения, т. к. перепад температур может вызвать темпера¬турный шок и повлиять на энергетический обмен и активность фер¬ментов. Так, параметры для пивных дрожжей указывают, что этот пе-репад не должен превышать 4°С.
Дрожжи, как и другие эукариоты, способны проявлять устойчи¬вость к тепловому шоку. Этот эффект основан на образовании це¬лого ряда так называемых протеинов теплового шока – «heat shock proteins». Приобретение термоустойчивости в результате шокового воздействия высоких температур обеспечивает также защиту дрож¬жей и от других воздействий. Например, увеличивается спирто- и осмоустойчивость.
Регулируя температуру, можно управлять микробиологическими процессами. Вместе с тем необходимо помнить, что низкие темпе¬ратуры легче переносятся большинством микроорганизмов, чем вы¬сокие. При снижении температуры вегетативные клетки переходят в анабиотическое состояние, при котором резко снижается их метабо¬лизм. Известны бактерии, остающиеся жизнеспособными при темпе¬ратуре ниже минус 190°С.
Высокая температура, как правило, губительно действует на веге¬тативные формы клеток и вирусные частицы в результате денатура¬ции белков. Действие высоких температур является одним из спосо¬бов повышения биологической стойкости напитков и других пищевых продуктов. К тепловым методам обработки пищи относятся стерили-зация и пастеризация.
Кислотность среды. Реакция среды, в которой находятся микроор¬ганизмы, оказывает на них большое влияние. Это один из наиболее важных факторов, определяющих рост и размножение микроорганиз¬мов, так как он определяет доступность для организма различных ве¬ществ и неорганических ионов.
Ниже показаны значения рН, в пределах которых возможен рост микроорганизмов:
| p. Bacillus | 4,5-8,5 |
| p. Lactobacillus | 3,0-6,5 |
| p. Acetobacter | 2,5-6,5 |
| p. Staphylococcus | 4,5-8,5 |
| p. Escherichia | 3,5-9,5 |
| p. Pseudomonas | 3,0-11,0 |
| p. Streptomyces | 4,5-8,5 |
| Дрожжи | 1,5-8,5 |
| Мицелиальные грибы | 1,5-9,0 |
Большинство микроорганизмов лучше всего растут при рН 7,0.
Величина рН влияет на диссоциацию кислот и оснований, следо¬вательно, оказывает влияние на перенос питательных веществ внутрь клетки, а также на степень токсичности ингибиторов роста. Недиссоциированные вещества, например органические кислоты, обла¬дают более высокой растворимостью в липидах, чем ионизированные формы, и поэтому снижение рН способствует большему проникно¬вению кислот в клетку. Также величина рН может воздействовать на конформацию молекул ферментов и изменять как первичный, так и вторичный метаболизм клетки. Очень кислая или очень щелочная реакция среды обычно токсична для бактерий. Поддержание опре¬деленного значения рН во время роста важно прежде всего для тех микроорганизмов, которые хотя и продуцируют кислоты, но сами не проявляют к ним толерантности (p. Lactobacillus, p. Enterobacteriaceae и многие представители p. Pseudomonas). Лишь некоторые виды бак-терий являются кислототолерантными (молочнокислые бактерии, Acetobacter, Sarcina) или даже ацидофильными (Thiobacillus, Acetobacter acidophilus).
Микроорганизмы, способные развиваться в очень кислой среде, объединяют в группу ацидофильных микроорганизмов. Они облигатно нуждаются для роста при очень низких значениях рН (3 и менее).
Многие микроорганизмы предпочитают слегка щелочную среду (нитрификаторы и клубеньковые бактерии, актиномицеты). Вместе с тем имеются представители, которые лучше растут при щелочной реакции (рН 10 и выше). Такие организмы называются алкалофилъными.
Свет. Прямые солнечные лучи обладают бактерицидной актив¬ностью. Такое действие света обусловлено активностью ультрафио¬летовой части спектра (длина волн 254-300 нм). Летальный эффект ультрафиолетовых лучей (УФ-лучи) с длиной волны около 260 нм объясняется тем, что именно в этой области лежит максимум погло¬щения молекул ДНК и РНК. Адсорбция УФ-лучей белками и нукле¬иновыми кислотами приводит к необратимым химическим измене¬ниям макромолекул. Происходит образование тиминовых димеров в ДНК бактериальных клеток. Сублетальные дозы УФ-лучей оказы-вают мутагенное действие на бактерии и вирусы. Наиболее чувстви¬тельны к действию света вегетативные клетки; для гибели спор необ¬ходимо в 4–5 раз больше энергии.
Бактерицидное действие ультрафиолета используют для обезза¬раживания вод, стерилизации воздуха в закрытых помещениях, для обеззараживания производственного оборудования. Обработка воз¬духа в течение 6 ч уничтожает до 80% микроорганизмов. Вместе с тем УФ-лучи обладают плохой проникающей способностью, поэтому не могут быть использованы для стерилизации пищевых продуктов.
Ультразвук, высокое давление, механическое воздействие. Ультра¬звук (УЗ) – это механические колебания с частотами свыше 20 000 Гц, что находится за пределами частот, воспринимаемых слухом человека. УЗ оказывает на клетки губительное действие. Это связано с образо¬ванием кавитационных полостей в цитоплазме бактерий, в которых создается высокое давление жидкостей, достигающее 10 000 атм, что приводит к разрушению цитоплазматических структур клетки. Споры бактерий более устойчивы, чем вегетативные клетки.
На жизнедеятельность микроорганизмов также оказывает влияние гидростатическое давление. При сбраживании сусла в высоких ЦКТ (высотой более 20 м) дрожжи испытывают гидростатический стресс, который усиливается растворенной углекислотой и возникающим давлением более 2 бар. При этом изменяются проницаемость дрож¬жевых клеток и их ферментативная активность.
На различных этапах пивоварения клетки испытывают и механи¬ческий стресс в результате действия больших касательных напряже¬ний, возникающих при перекачке и центрифугировании, движении по трубопроводу.
Обработка дрожжей и реакция клеток на излишнее механиче¬ское воздействие приводят к возникновению таких явлений, как истощение клеточной стенки, выделение внутриклеточных суб¬станций.
«Классическая» реакция дрожжевой клетки на механический стресс – это снижение уровня гликогена и ее жизнеспособности, увеличение рН, модификация трегалозы, выход из клетки внут¬риклеточной протеазы. Менее классическим проявлением влияния стресса является изменение функциональных свойств клеточной стенки. С увеличением времени воздействия стресс-факторов про¬исходит выход инвертазы (фермента, ассоциированного с клеточной стенкой), а также маннана и глюкана на поверхность, что говорит об изношенности стенок клеток в результате трения во время механи¬ческого воздействия.
Таким образом, вполне закономерными последствиями изношен¬ности клеточных стенок являются нарушение флокуляционной спо¬собности и уменьшение количества клеток в суспензии во время фер¬ментации. Такая реакция клетки на стресс приводит к изменениям в процессе флокуляции, плохой фильтрации, низкой пеностойкости и в конечном итоге к некачественному пиву.
Возникающие в трубопроводах турбулентные и ламинарные потоки также оказывают воздействие на клетки. В целом, турбулентное дви¬жение считается энергетически более насыщенным и соответственно более опасным для клеток. Однако было показано, что ламинарное движение в отличие от турбулентного вызывает большую реакцию со стороны клетки, о чем говорят увеличение выхода протеазы и сни¬жение уровня ферментации. Такое поведение объясняется взаимо¬связью между временем пребывания в пристеночной области (где уровень стресса наиболее высокий) и средней скоростью воздействия в этой области (она ниже в случае ламинарного потока). Хотя воз¬действие на околостенную область выше при турбулентном потоке, время пребывания клеточки в этой области при том же самом движе¬нии меньше и, таким образом, необходимо минимум времени на реа¬гирование клетки.
Влияние кислорода. По отношению к молекулярному кислороду выделяют несколько групп микроорганизмов.
Облигатные, или строгие, аэробы. Они могут жить и размножаться только в присутствии кислорода, т. к. используют его в качестве ко¬нечного акцептора электронов и протонов в процессе окислительного фосфорилирования при аэробном дыхании.
Для бактерий, растущих на твердой питательной среде в присут¬ствии воздуха, кислорода обычно достаточно. В жидких средах аэроб¬ные микроорганизмы растут только на поверхности, т. к. по мере удаления от поверхности условия приближаются к анаэробным. Для роста аэробов в жидких питательных средах необходимо проводить постоянную аэрацию среды. Представителями аэробных микроорга¬низмов являются представители р. Bacillus, уксуснокислые бактерии, многие актиномицеты, мицелиальные грибы.
Строгие аэробы могут развиваться в присутствии кислорода, т. к. у них есть ферменты каталаза и супероксиддисмутаза (СОД), защи¬щающие клетку от токсического действия образующихся в ходе био¬химических реакций свободных радикалов, например, супероксиданиона (• О2-) и других реакционно-способных кислородсодержащих соединений, в частности Н2О2.
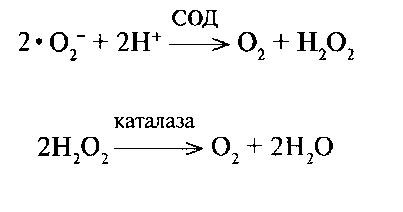
Микроаэрофилы, или факультативные аэробы. Они развиваются и при незначительной концентрации кислорода в среде (2-5%). Такой способностью обладают бактерии – представители родов Micrococcus, Campylobacter.
Облигатные анаэробы. У анаэробных микроорганизмов конструк¬тивные и энергетические процессы в клетке протекают без участия кислорода. Для многих он просто яд. При анаэробном дыхании ко¬нечным акцептором водорода является не кислород, а чаще всего окисленные соединения азота (нитраты, нитриты), соединения серы (сульфаты, сульфиты, тиосульфаты) или окисленные соединения уг¬лерода (диоксид углерода, бикарбонаты).
У анаэробных микроорганизмов очень широко распространен процесс брожения как способ получения энергии. При брожении ко¬нечным акцептором электронов и протонов, полученных в ходе мета¬болизма одного органического вещества, служит другое органическое вещество-метаболит. Чаще всего это пировиноградная кислота (ПВК) или продукт ее превращения. Перенос водорода от доноров к акцеп¬торам осуществляется с помощью окислительно-восстановительных реакций. Все брожения, проводимые микроорганизмами, имеют общие черты: они энергетически низко эффективны, окислению под¬вергается большое количество органического субстрата, образуется большое количество конечных продуктов метаболизма и малый прирост биомассы.
Факультативные анаэробы. Представители этой группы микроор¬ганизмов могут расти и размножаться как в присутствии кислорода, так и при его малой концентрации в среде. Факультативными анаэро¬бами являются бактерии семейства Enterobacteriaceae, представители p. Staphylococcus, дрожжи p. Saccharomyces.
В мире микроорганизмов это уникальные организмы, т. к. они способны переключать свой метаболизм с аэробного дыхания (окис¬лительного фосфорилирования) на брожение (субстратное фосфорилирование) в зависимости от условий их существования. Клетки в аэробных условиях получают значительно больше энергии в виде молекул АТФ и интенсивно накапливают биомассу. Являясь анаэробами, клетки дрожжей эволюционировали, выработали механизмы защиты от окислительного действия кислорода и смогли приспосо¬биться к жизни в аэробных условиях.
Способности дрожжей противостоять окислительному стрессу посвящено большое число исследований. В производстве пива созна¬тельная подача кислорода осуществляется только однажды – при аэ¬рации сусла, и стресс, вызванный кислородом, маловероятен, так как клетки подвергаются воздействию кислорода только в начале броже¬ния, к тому же его концентрация в среде невелика (менее 15 мг/дм3). Растворенный кислород потребляется дрожжами в течение всего не¬скольких часов, поэтому в ходе реакций, протекающих в митохонд¬риях, в клетки поступает небольшое количество активного кислорода, способного отрицательно влиять на жизнедеятельность дрожжей. Кроме того, необходимо отметить, что у клеток имеются свои защит¬ные механизмы – это ферменты: каталаза, супероксиддисмутаза, глутатионпероксидаза, тиоредоксинпироксидаза, метионинредуктаза, редуктаза, а также соединения: глутатион, полиамины, ионы металлов, тиоредоксин и др.
В таких условиях кислород рассматривается как субстрат, необ¬ходимый для синтеза ненасыщенных жирных кислот и эргостерина, которые нужны для роста клеток. Данная процедура направлена на улучшение физиологического состояния дрожжей и получение боль¬шей биомассы.
Углекислый газ. Различные газы, растворенные в среде, могут также влиять на жизнедеятельность микроорганизмов, и в частности дрожжей.
При росте и размножении дрожжевых клеток независимо от типа энергетического обмена (брожение или дыхание) образуется диоксид углерода. Растворенный в культуральной жидкости диоксид уг¬лерода угнетает рост дрожжей. Токсичное действие СО2 на дрожжи известно давно. При давлении газа около 0,5 атм цикл трикарбоновых кислот ингибируется, а спиртовое брожение продолжается вплоть до давления 4,0 атм. Деление клеток прекращается при давлении 2,5-3,0 атм. Интересно, что при этом они проходят S-фазу роста и имеют удвоенное содержание ДНК, и поэтому размер кле¬ток больше, чем обычно. Механизм действия СО2 очень разнороден и точно не установлен.
Растворимость диоксида углерода зависит от температуры среды и давления; так, при атмосферном давлении при 30°С в 1 дм3 воды рас¬творяется 1,2 г СО2 Избыток диоксида углерода в среде устраняется аэрированием культуральной жидкости.